Зміст
- Генетична інформація у прокаріотів
- Етапи експресії генів
- Транскрипція в бактеріальні клітини
- Транскрипція: етап ініціації
- Транскрипція: Фаза подовження
- Транскрипція: фаза припинення
- Переклад у клітини бактерій
- Переклад: Посвячення
- Переклад: Подовження
- Переклад: Припинення
- Переклад та антибіотики
- Обробка протеїну після перекладу
- Фосфорилювання білка
- Ацетилювання білка та глікозилювання
- Експресія генів в Археях
Прокаріоти - це дрібні одноклітинні живі організми. Вони є одним із двох поширених типів клітин: прокаріотичний і еукаріотичний.
Оскільки клітини прокаріотів не мають ядра або органел, експресія гена відбувається у відкритій цитоплазмі і всі стадії можуть відбуватися одночасно. Хоча прокаріоти простіші, ніж еукаріоти, контроль експресії генів все ще має вирішальне значення для їх клітинної поведінки.
Генетична інформація у прокаріотів
Два домені прокаріотів - бактерії та археї. Обидва не мають визначеного ядра, але вони все ще мають генетичний код і нуклеїнові кислоти. Хоча таких складних хромосом немає, як ви б побачили в еукаріотичних клітинах, прокаріоти мають кругові шматочки дезоксирибонуклеїнової кислоти (ДНК), розташовані в нуклеоїді.
Однак навколо генетичного матеріалу немає мембрани. Як правило, прокаріоти мають менше послідовностей, що не кодують, у своїй ДНК порівняно з еукаріотами. Це може бути пов'язано з тим, що прокаріотичні клітини мають менші розміри та мають менше місця для молекули ДНК.
The нуклеоїд просто область, де ДНК живе в клітині прокаріотів. Він має неправильну форму і може відрізнятися розмірами. Крім того, нуклеоїд прикріплюється до клітинної мембрани.
Прокаріоти також можуть мати кругову ДНК плазміди. У них можливо одна або кілька плазмід у клітині. Під час поділу клітин прокаріоти можуть проходити шляхом синтезу ДНК та поділу плазмід.
Порівняно з хромосомами еукаріотів, плазміди мають менший розмір і мають менше ДНК. Крім того, плазміди можуть самостійно реплікуватися без іншої клітинної ДНК. Деякі плазміди несуть коди несуттєвих генів, наприклад, такі, що надають бактеріям антибіотикорезистентність.
У певних випадках плазміди також здатні переходити з однієї клітини в іншу клітину та обмінюватися інформацією, такою як стійкість до антибіотиків.
Етапи експресії генів
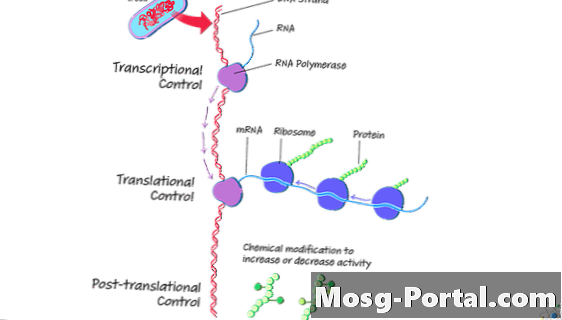
Експресія гена - це процес, за допомогою якого клітина перетворює генетичний код в амінокислоти для отримання білка. На відміну від еукаріотів, дві основні стадії, які є транскрипцією та трансляцією, можуть відбуватися одночасно у прокаріотів.
Під час транскрипції клітина переводить ДНК в молекулу месенджерної РНК (мРНК). Під час трансляції клітина виробляє амінокислоти з мРНК. Амінокислоти становлять білки.
І транскрипція, і трансляція відбуваються у прокаріотів цитоплазма. За умови, що обидва процеси відбуваються одночасно, клітина може виробляти велику кількість білка з одного і того ж шаблону ДНК. Якщо клітині білок більше не потрібен, транскрипція може припинитися.
Транскрипція в бактеріальні клітини
Метою транскрипції є створення комплементарного ланцюга рибонуклеїнової кислоти (РНК) із шаблону ДНК. Процес складається з трьох частин: ініціювання, подовження ланцюга та припинення.
Для того, щоб відбулася фаза ініціації, ДНК повинна спочатку розмотатися, і область, де це відбувається, - це міхур транскрипції.
У бактеріях ви знайдете ту саму РНК-полімеразу, відповідальну за всю транскрипцію. Цей фермент має чотири субодиниці. На відміну від еукаріотів, прокаріоти не мають факторів транскрипції.
Транскрипція: етап ініціації
Транскрипція починається, коли ДНК розмотується і РНК-полімераза зв'язується з промотором. Промотор - це особлива послідовність ДНК, яка існує на початку конкретного гена.
У бактерій промотор має дві послідовності: -10 і -35 елементів. Елемент -10 - це місце, де ДНК зазвичай розмотується, і в ньому розташовано 10 нуклеотидів від місця ініціації. Елемент -35 - це 35 нуклеотидів з сайту.
РНК-полімераза спирається на один ланцюг ДНК, який є шаблоном, оскільки він будує нову ланцюг РНК, що називається транскриптом РНК. Отриманий ланцюг РНК або первинний стенограф майже такий же, як нешаблон або кодує ланцюг ДНК. Різниця полягає лише в тому, що всі основи тиміну (Т) є основами урацилу (U) в РНК.
Транскрипція: Фаза подовження
Під час фази транскрипції ланцюга подовження ланцюга РНК-полімераза рухається вздовж ланцюга шаблону ДНК і утворює молекулу мРНК. Нитка РНК стає довшою, оскільки додається більше нуклеотидів.
По суті, РНК-полімераза проходить уздовж ДНК-стенда у напрямку від 3 до 5 для цього. Важливо зазначити, що бактерії можуть створювати полікістронні мРНК що код для кількох білків.
••• Наукові роботиТранскрипція: фаза припинення
Під час фази припинення транскрипції процес зупиняється. У прокаріотів існує два типи фаз термінації: Rho-залежне закінчення та Rho-незалежне припинення.
В Розозалежне припинення, спеціальний білковий фактор під назвою Rho перериває транскрипцію і припиняє її. Протеїновий фактор Rho приєднується до ланцюга РНК на конкретному місці зв'язування. Потім він рухається по ланцюжку, щоб досягти РНК-полімерази в міхурі транскрипції.
Далі, Rho роз'єднує нову ланцюг РНК та шаблон ДНК, тому транскрипція закінчується. Полімераза РНК перестає рухатися, оскільки вона досягає кодуючої послідовності, яка є точкою зупинки транскрипції.
В Rho-незалежне припинення, молекула РНК робить петлю і від'єднується. РНК-полімераза досягає послідовності ДНК на нитці шаблону, яка є термінатором і має багато нуклеотидів цитозину (С) та гуаніну (G). Нова пасмо РНК починає складатись у форму шпильки. Його нуклеотиди C і G зв'язуються. Цей процес зупиняє рух РНК-полімерази.
Переклад у клітини бактерій
Переклад створює молекулу білка або поліпептид на основі шаблону РНК, створеного під час транскрипції. У бактерій переклад може статися відразу, а іноді він починається під час транскрипції. Це можливо через те, що у прокаріотів немає ядерних мембран і жодних органел для поділу процесів.
У еукаріотів речі різні, тому що транскрипція відбувається в ядрі, а трансляція - у цитозолабо внутрішньоклітинна рідина клітини. Еукаріот також використовує зрілу мРНК, яку обробляють перед трансляцією.
Ще одна причина, чому трансляція та транскрипція можуть відбуватися одночасно у бактерій, - це те, що РНК не потребує спеціальної обробки, що спостерігається у еукаріотів. РНК бактерій готовий до перекладу негайно.
Смуга мРНК має групи нуклеотидів, які називаються кодони. Кожен кодон має три нуклеотиди та коди для певної амінокислотної послідовності. Хоча є лише 20 амінокислот, клітини мають 61 кодон для амінокислот і три стоп-кодони. AUG є початковим кодоном і починає переклад. Він також кодує амінокислоту метіонін.
Переклад: Посвячення
Під час трансляції нитка мРНК виступає в якості шаблону для отримання амінокислот, які стають білками. Клітина декодує мРНК для цього.
Ініціація вимагає передачі РНК (тРНК), рибосоми та мРНК. Кожна молекула тРНК має антикодон для амінокислоти. Антикодон доповнює кодон. У бактерій процес починається, коли невелика рибосомальна одиниця приєднується до мРНК при a Послідовність блиску-далгарно.
Послідовність Shine-Dalgarno є особливою рибосомальною зоною зв'язування як бактерій, так і архей. Зазвичай ви бачите це приблизно вісім нуклеотидів від стартового кодона AUG.
Оскільки бактеріальні гени можуть мати транскрипцію у групах, одна мРНК може кодувати багато генів. Послідовність Shine-Dalgarno полегшує пошук стартового кодона.
Переклад: Подовження
Під час подовження ланцюг амінокислот стає довшим. ТРНК додають амінокислоти для створення поліпептидного ланцюга. ТРНК починає працювати в P сайт, що є середньою частиною рибосоми.
Поруч із сайтом Р розташований сайт Сайт. ТРНК, що відповідає кодону, може перейти на сайт А. Тоді між амінокислотами може утворюватися пептидна зв’язок. Рибосома рухається вздовж мРНК, а амінокислоти утворюють ланцюг.
Переклад: Припинення
Припинення відбувається через стоп-кодон. Коли стоп-кодон потрапляє на сайт А, процес трансляції припиняється, оскільки стоп-кодон не має комплементарної тРНК. Білки називають фактори вивільнення які поміщаються в P-ділянку, можуть розпізнати стоп-кодони і запобігти утворенню пептидних зв'язків.
Це відбувається тому, що фактори вивільнення можуть змусити ферменти додавати молекулу води, завдяки чому ланцюг відокремлюється від тРНК.
Переклад та антибіотики
Якщо ви приймаєте деякі антибіотики для лікування інфекції, вони можуть працювати, порушуючи процес перекладу бактерій. Мета антибіотиків - вбити бактерії та зупинити їх розмноження.
Один із способів цього досягти - це вплинути на рибосоми в бактеріальних клітинах. Препарати можуть перешкоджати трансляції мРНК або блокувати здатність клітини здійснювати пептидні зв’язки. Антибіотики можуть зв’язуватися з рибосомами.
Наприклад, один тип антибіотика під назвою тетрациклін може потрапити в бактеріальну клітину шляхом перетину плазматичної мембрани і накопичення всередині цитоплазми. Тоді антибіотик може зв’язуватися з рибосомою і блокувати трансляцію.
Інший антибіотик під назвою ципрофлоксацин впливає на бактеріальні клітини, націлюючи фермент, відповідальний за розмотування ДНК, щоб дозволити реплікацію. В обох випадках клітини людини шкодуються, що дозволяє людям вживати антибіотики, не вбиваючи власні клітини.
Пов'язана тема: багатоклітинні організми
Обробка протеїну після перекладу
Після закінчення трансляції деякі клітини продовжують обробку білків. Пост-трансляційні модифікації (PTM) білків дозволяють бактеріям адаптуватися до свого середовища та контролювати поведінку клітин.
Взагалі ПТМ зустрічаються рідше у прокаріотів, ніж еукаріоти, але деякі організми мають їх. Бактерії також можуть змінювати білки і змінювати процеси. Це надає їм більшої універсальності та дозволяє використовувати модифікацію білка для регулювання.
Фосфорилювання білка
Фосфорилювання білка є поширеною модифікацією бактерій. Цей процес включає додавання фосфатної групи до білка, в якому є атоми фосфору та кисню. Фосфорилювання має важливе значення для функції білка.
Однак фосфорилювання може бути тимчасовим, оскільки воно є оборотним. Деякі бактерії можуть використовувати фосфорилювання як частину процесу для зараження інших організмів.
Викликається фосфорилювання, яке відбувається в бічних ланцюгах серинної, треонінової та тирозинової амінокислот Фосфорилювання Ser / Thr / Tyr.
Ацетилювання білка та глікозилювання
Крім фосфорильованих білків, бактерії можуть мати ацетильований і глікозильований білки. Вони також можуть мати метилювання, карбоксилювання та інші модифікації. Ці модифікації відіграють важливу роль у сигналізації, регуляції клітин та інших процесах бактерій.
Наприклад, фосфорилювання Ser / Thr / Tyr допомагає бактеріям реагувати на зміни свого середовища та збільшувати шанси на виживання.
Дослідження показують, що метаболічні зміни в клітині пов'язані з фосфорилюванням Ser / Thr / Tyr, що вказує на те, що бактерії можуть реагувати на їхнє середовище, змінюючи свої клітинні процеси. Більше того, пост-трансляційні модифікації допомагають їм швидко та ефективно реагувати. Можливість зміни будь-яких змін також забезпечує суттєвий контроль.
Експресія генів в Археях
Археї використовують механізми експресії генів, більш схожі на еукаріоти. Хоча археї є прокаріоти, вони мають деякі спільні з еукаріоти, такі як експресія генів та регуляція генів. Процеси транскрипції та трансляції в археях також мають певну схожість з бактеріями.
Наприклад, і археї, і бактерії містять метіонін в якості першої амінокислоти і AUG як вихідний кодон. З іншого боку, і археї, і еукаріоти мають а Коробка TATA, що являє собою послідовність ДНК в області промотору, яка показує, де розшифрувати ДНК.
Переклад у археї нагадує процес, який спостерігають у бактерій. Обидва типи організмів мають рибосоми, які складаються з двох одиниць: субодиниці 30S та 50S. Крім того, обидва вони мають полікістронні мРНК та послідовності Shine-Dalgarno.
Між бактеріями, археями та еукаріотами існує багато подібностей та відмінностей. Однак всі вони покладаються на експресію генів та регуляцію генів, щоб вижити.